
The term fluid applies to both liquids and gases. The essential differences between fluids and solids can be summarized as follows:
1 P = 0.1 kg / (m s).It is most often described in terms of cgs units: units whose fundamental measures are the gram, the centimeter and the second. In cgs units, since there are 1000 grams in a kilogram and 100 centimeters in a meter, we have
1 P = 0.1 kg / (m s) * (1000 g / kg) * (1 m / 100 cm)The viscosity of water is .01 Poise, and the viscosity of blood is .04 Poise.= 1 g / (cm s).
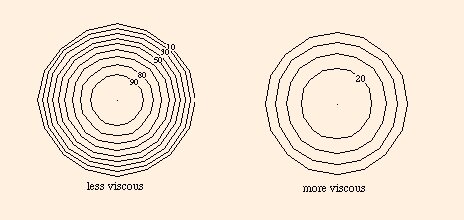
the velocity does not depend on the angular position on any concentric circle, but only on the distance from the center.
The fact that the velocity is constant on each circle leads us to think of the fluid as flowing in concentric sheets, but in fact the velocity is a smooth function of distance from the center. For higher viscosity, the velocity gradient is shallower. Essentially, viscosity opposes the existence of steep velocity gradients. The larger the viscosity, the gentler is the shape of the parabolic velocity gradient, and the velocity is more nearly constant across the section.
Fluid flow in a pipe crosses the threshold from laminar to turbulent flow when a dimensionless parameter called the Reynold's Number
(denoted Re) reaches about 2000. It is defined as
Re = 2 ρ r v / η,
where ρ denotes the density of the fluid (1.05 g / cm3 for blood), v is the fluid's velocity
and η is the Greek letter eta denoting the viscosity. Re is essentially the ratio of the inertial
forces (tending to keep the fluid flowing) to the viscous forces experienced by a layer of fluid. Its value indicates the relative
unimportance of viscosity (ie., low Re corresponds to very viscous situations). Typical values for human arteries and veins are:
vessel r v Re artery .2 cm 28 cm/s 294 vein .25 cm 36 cm/s 472.5
For laminar flow through a uniform straight pipe, the flow rate (volume per unit time) is given by Poiseuille's Equation:
We would like to use Poiseuille's Equation to discuss the blood vessels in animal circulatory systems. While the flow is essentially
laminar outside of the capillaries, it is pulsatile throughout the arterial subsystem: the pressure varies as a periodic function of time.
In addition, the equation is based on the parabolic velocity gradient, but since pressure waves in arterial walls propagate more
quickly than those in blood, the velocity profile is more uniform than parabolic. Beyond that, Poiseuille's Equation assumes a
constant viscosity, whereas the viscosity of blood actually changes with velocity, since blood is not a uniform fluid. In fact, the
viscosity is much lower in the capillaries than in the rest of the system, since the red blood cells line up in single file to pass
through them. On top of everything else, the blood vessels are not straight, uniform pipes!
All of these reservations notwithstanding, we can apply Poiseuille's Equation to the circulatory system to understand the scaling
relationships between the various parameters. For instance, an occluded artery is often the subject of angioplasty or even bypass,
in order to increase the inner radius of the vessel. We can use Poisseuille's Equation to understand the functional dependency of
flow on radius: since flow is proportional to the fourth power of radius, we see that increasing the radius by a factor of 2 results
in an increase in the flow rate by a factor of 16. We say that the flow scales with the fourth power of the radius. In a similar
fashion, we can relate any two of the variables, holding all of the others constant. Since flow is equal to velocity times cross
sectional area, we can also relate the blood velocity and therefore the Reynold's number to the other parameters.
It is necessary to have a single equation which contains exactly two interrelated variables in order to deduce a scaling relationship.
In the event that three variables are interrelated through two or more equations, one variable must be eliminated. Any single equation
which contains three interrelated variables is insufficient to deduce the relation; in that case a different equation is necessary.
Note that in this applet, the answers must be exact! Remember that answers less than one must be entered as fractions (ie., 1/2).
The hydrostatic pressure in a volume of fluid is the pressure at any given depth (d) due to the weight of the fluid above that depth:
Friction often presents itself as drag when an object is moving through a fluid medium.
It is a force proportional to the object's velocity, but opposite to the direction of motion:
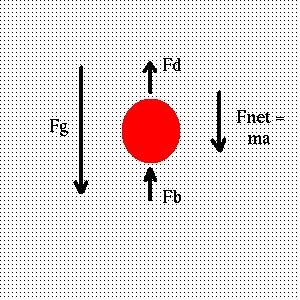
Consider an object sinking in a swimming pool:
Opposing gravity is the buoyant force of the water, which attempts to raise the object, and the drag force, which slows
its sinking. The buoyant force is equal to the weight of the fluid displaced by the object:
Equating the sum of the forces due to gravity, buoyancy and drag to the mass times the acceleration, and taking both V and A to be
positive for a sinking object, we find
In electrophoresis, which is used in DNA analysis and the determination of protein molecular weights,
an electric field takes the place of gravity and buoyancy. The acceleration equation becomes
The next section builds an electrical analogy to fluid systems.
©2009, Kenneth R. Koehler. All Rights Reserved. This document may be freely reproduced provided that this copyright notice is included.
Please send comments or suggestions to the author.
F = ΔP π r 4 / 8 η l,
where ΔP is the pressure drop experienced by the fluid as a result of viscous losses along the length l of the pipe.
P = ρ g d
This pressure is independent of any atmospheric or surface pressure exerted at depth d = 0.
. . .
Fd = - b V.
Here b is the drag coefficient, with dimensions of mass over time. The drag coefficient depends on the radius and shape of the
object, and the viscosity of the medium. For a sphere of radius r, the drag coefficient is
b0 = 6 π η r.
For other shapes, we multiply by a shape factor b / b 0 which depends on the shape of the object;
for a more cylindrical object, the shape factor might be around 1.5.
Fb = m v ρ g.
Here v is the specific volume of the object (volume per unit mass).
Thus m v gives the volume of the object, and ρ g gives the gravitational force per unit volume of the fluid.
m g - m v ρ g - b V = m A
or
(1 - v ρ) m g - b V = m A
First note that the object floats if v ρ > 1, that is, if the density of the object is less than that of the
fluid. If the object does sink, V is initially small, A is positive and V therefore grows. But A clearly decreases, and at
some point becomes zero. At that point, V stays constant. The exact result requires calculus, but
for large times relative to m / b, the velocity approaches the terminal velocity:
Vt = (1 - v ρ) m g / b.
Note that the hydrostatic pressure does not affect this result, since it exerts a force on the object equally in both the
upward and downward directions.
q E - b V = m A
and the terminal velocity is then
Vt = q E / b
The molecules rapidly reach terminal velocity, and the measurement of the velocity (or correspondingly, the distance the molecules
travel in a given time) determines the charge on the molecule. Provided that the molecules have been denatured (unfolded)
and charged by the addition of an anionic detergent like sodium dodecylsulfate (SDS), the charge will be proportional to the
mass of the molecule and the molecular weight can be determined.